Sintesi resoconto 2015 - Gruppo Inanellamento Limicoli (GIL, Napoli)
Menu principale:
- Home Page
- L'Associazione
-
Ricerche in atto
- Migrazione autunnale Allodola
- Migrazione primaverile Tordo
- Ali Tordo bottaccio in Campania
- Stazione Monte Vaccaro
- Riserva Naturale Foce Volturno
- Inanellamento Medio Volturno
- Avifauna svernante
- Avifauna Foce Neto e Sila
-
Ricerche concluse
- Ali Turdidi e Beccacce ATC LT 2
- Stazione Malga Rosello
- Migrazione Caradriiformi
- Progetto Rondine
- Migrazione autunnale Tordo
- Ali Turdidi e Beccacce ATC CE
- Progetto Alauda 2000
- Osservatorio Roccolo Ganda
- Atlante Migrazioni Puglia
- Download
- News
Sintesi resoconto 2015
Studio sulla migrazione autunnale dell'Allodola in Campania: sintesi dei dati autunno 2015
Gruppo Inanellamento Limicoli (G.I.L., Napoli)
Federazione Italiana della Caccia -
Gli studi sulle tendenze demografiche a lungo termine e sull’andamento della migrazione autunnale di questa specie sono piuttosto scarsi; l'unica eccezione è rappresentata dal progetto "Alouette", organizzato dal 1990 in Francia dal “Centre de Recherches sur la Biologie des Populations d'Oiseaux”, e tutt’ora in corso con più di 58.000 uccelli inanellati (Teulière 2003). L'analisi dei dati raccolti ha permesso di quantificare il flusso migratorio durante la migrazione post-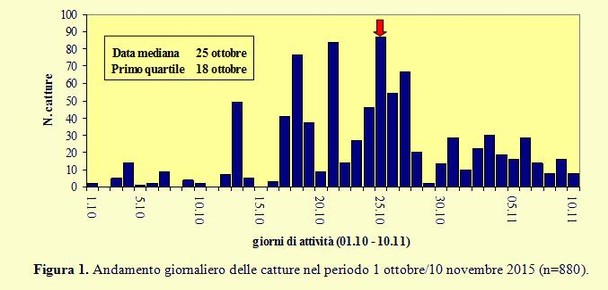 Il grafico mostra un numero di catture estremamente ridotto durante tutta la prima decade di ottobre, in cui si sono avuti venti per lo più provenienti dai quadranti meridionale ed occidentale e condizioni di tempo perturbato. Anche la seconda decade di ottobre è stata caratterizzata dalle medesime condizioni atmosferiche ed ha fatto registrare numeri piuttosto bassi di catture con solo quattro giornate in cui sono stati inanellati più di 10 individui e con un picco di 77 catture il 18 ottobre. Nella terza decade di ottobre, pur permanendo condizioni meteorologiche instabili, si sono registrati per lo più venti settentrionali accompagnati da un maggiore flusso migratorio che ha consentito catture più numerose. In nessuna giornata è stata superata la soglia delle 100 catture ed il picco massimo è stato di 87 inanellati il 25 ottobre. Nei giorni successivi e fino alla fine del periodo di monitoraggio i passaggi sono sempre stati molto modesti e di conseguenza anche il numero delle catture, sebbene le condizioni meteo registrate nell’area siano migliorate con venti provenienti dai quadranti settentrionali. In base alla distribuzione temporale delle catture la data mediana (quando cioè è stato catturato il 50% di tutti gli uccelli), evidenziata nel grafico dalla freccia, è caduta il 25 ottobre mostrando una notevole variabilità nel corso degli anni: rispetto a quella registrata nello scorso anno è avvenuta con 11 giorni di ritardo, sei rispetto al 2012 e cinque rispetto al 2013, mentre è risultata in anticipo di tre giorni rispetto al 2011; il 18 ottobre è terminato il primo quartile ed il 28 ottobre il terzo quartile: tra queste due date è risultato compreso il periodo principale della migrazione.
Il grafico mostra un numero di catture estremamente ridotto durante tutta la prima decade di ottobre, in cui si sono avuti venti per lo più provenienti dai quadranti meridionale ed occidentale e condizioni di tempo perturbato. Anche la seconda decade di ottobre è stata caratterizzata dalle medesime condizioni atmosferiche ed ha fatto registrare numeri piuttosto bassi di catture con solo quattro giornate in cui sono stati inanellati più di 10 individui e con un picco di 77 catture il 18 ottobre. Nella terza decade di ottobre, pur permanendo condizioni meteorologiche instabili, si sono registrati per lo più venti settentrionali accompagnati da un maggiore flusso migratorio che ha consentito catture più numerose. In nessuna giornata è stata superata la soglia delle 100 catture ed il picco massimo è stato di 87 inanellati il 25 ottobre. Nei giorni successivi e fino alla fine del periodo di monitoraggio i passaggi sono sempre stati molto modesti e di conseguenza anche il numero delle catture, sebbene le condizioni meteo registrate nell’area siano migliorate con venti provenienti dai quadranti settentrionali. In base alla distribuzione temporale delle catture la data mediana (quando cioè è stato catturato il 50% di tutti gli uccelli), evidenziata nel grafico dalla freccia, è caduta il 25 ottobre mostrando una notevole variabilità nel corso degli anni: rispetto a quella registrata nello scorso anno è avvenuta con 11 giorni di ritardo, sei rispetto al 2012 e cinque rispetto al 2013, mentre è risultata in anticipo di tre giorni rispetto al 2011; il 18 ottobre è terminato il primo quartile ed il 28 ottobre il terzo quartile: tra queste due date è risultato compreso il periodo principale della migrazione.
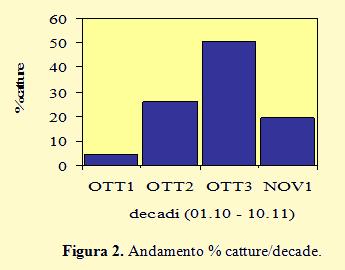 Nella figura 2 la distribuzione delle catture è organizzata per decade. Diversamente dai due anni precedenti, la percentuale più alta è stata ottenuta nella terza decade (50,2% con 442 inanellati) con un discreto valore anche nella seconda (25,9% con 228 catture), mentre la prima decade ha fatto registrare la percentuale più bassa (4,4%, solo 39 inanellati) ottenuta durante il periodo di studio.
Nella figura 2 la distribuzione delle catture è organizzata per decade. Diversamente dai due anni precedenti, la percentuale più alta è stata ottenuta nella terza decade (50,2% con 442 inanellati) con un discreto valore anche nella seconda (25,9% con 228 catture), mentre la prima decade ha fatto registrare la percentuale più bassa (4,4%, solo 39 inanellati) ottenuta durante il periodo di studio.
Nella tabella 1 si osserva come i numeri relativi alle catture effettuate nelle quattro decadi di questa stagione risultano marcatamente più bassi di quelli registrati negli anni precedenti con una drastica flessione nella prima decade di ottobre ed un numero complessivo di inanellati ancora inferiore a quello già basso dell’anno precedente che corrisponde ad un flusso migratorio rimasto modesto per tutta la stagione.
Come negli anni precedenti, anche se con evidenti fluttuazioni nel numero degli inanellati da un anno all’altro, anche nel 2015 il maggior numero di inanellati è concentrato nella seconda e nella terza decade di ottobre, mentre nella prima decade di novembre le catture sono diminuite notevolmente. Si può osservare inoltre come in alcune annate, così come quest’anno, il numero di inanellati della prima decade di ottobre risulti davvero modesto.
OTT 1 |
OTT 2 |
OTT 3 |
NOV 1 |
TOT |
|
2006 |
421 |
566 |
494 |
81 |
1.562 |
2007 |
322 |
419 |
589 |
173 |
1.503 |
2010 |
34 |
201 |
1.142 |
193 |
1.570 |
2011 |
77 |
335 |
750 |
269 |
1.431 |
2012 |
399 |
1.169 |
300 |
231 |
2.099 |
2013 |
114 |
523 |
426 |
89 |
1.152 |
2014 |
252 |
468 |
120 |
135 |
975 |
2015 |
39 |
228 |
442 |
171 |
880 |
1.658 |
3.909 |
4.263 |
1.342 |
11.172 |
|
Tabella 1. Numero di inanellati per decade (dati 2006 e 2007 da Scebba 2009). |
|||||

L'utilizzo di una metodologia di cattura completamente standardizzata consente di calcolare un indice di cattura (IC) con cui è possibile valutare con maggior precisione l’andamento dei passaggi in quanto il numero di uccelli catturati è rapportato al numero di ore in cui le reti sono rimaste operative. Viene quindi considerata tutta l’attività espletata, dalla prima sessione di cattura effettuata nel mese di ottobre fino all’ultima avvenuta in novembre, in modo da coprire l’intero periodo della migrazione (tabella 2). L’analisi dei dati raccolti in questi otto anni mostra che l’indice di cattura, rimasto sostanzialmente costante per gli anni 2006, 2007, 2010 e 2011, ha fatto registrare un deciso incremento nel 2012 mentre a partire dal 2013 presenta una flessione notevole che in questo ultimo anno di studio risulta ancora più marcata.
Anno |
n° catture |
mqrete x ore |
IC |
2006 |
1.562 |
73260 |
21,32 |
2007 |
1.503 |
72600 |
20,70 |
2010 |
1.570 |
73260 |
21,43 |
2011 |
1.454 |
68640 |
21,18 |
2012 |
2.099 |
70620 |
29,68 |
2013 |
1.152 |
65340 |
17,63 |
2014 |
975 |
69960 |
13,94 |
2015 |
880 |
73260 |
12,01 |
Tabella 2. Indice di Cattura (IC) per stagione |
|||
Il campione ottenuto nel 2015 è risultato composto per il 4,9% (43 uccelli) da soggetti il cui sesso resta indeterminato, per il 32,8% (289) da maschi e per il 62,3% (548) da femmine, percentuali che non si discostano significativamente da quelle ottenute nello scorso anno. Calcolando la sex-

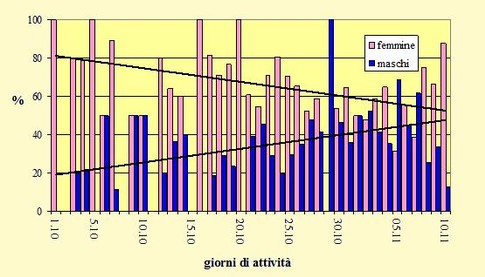
Figura 3. Fenologia della migrazione autunnale basata sulla percentuale di maschi e femmine nel 2015 con linee di tendenza.
In base alla distribuzione temporale delle catture la data mediana è caduta il 24 ottobre per le femmine ed il 26 ottobre per i maschi; il primo quartile è terminato, rispettivamente il 18 ed il 21 ottobre, ed il terzo quartile il 27 ottobre ed il 1 novembre. Il periodo principale della migrazione, che include il 50% degli uccelli catturati, è quindi ricaduto tra il 18 ed il 27 ottobre per le femmine e tra il 21 ottobre ed il 1 novembre per i maschi.
L'esame dell’accumulo del grasso sottocutaneo rappresenta un aspetto importante per determinare lo stato fisiologico degli uccelli e valutare le strategie di migrazione adottate per raggiungere le aree di svernamento, in quanto tale deposito fornisce l’energia necessaria per sostenere la migrazione venendo consumato durante il volo. Nella tabella 3 sono riportate, per gli ultimi sei anni di studio, percentuali e numero di individui, separati per sesso, inclusi nelle diverse classi di grasso (Kaiser 1993).
Autunno 2010 |
Autunno 2011 |
Autunno 2012 |
Autunno 2013 |
Autunno 2014 |
Autunno 2015 |
|||||||
Classi di |
♂♂ |
♀♀ |
♂♂ |
♀♀ |
♂♂ |
♀♀ |
♂♂ |
♀♀ |
♂♂ |
♀♀ |
♂♂ |
♀♀ |
0 |
2 |
5 |
10 |
14 |
2 |
9 |
11 |
34 |
2 |
10 |
11 |
18 |
1 - |
162 |
297 |
86 |
203 |
96 |
370 |
145 |
391 |
49 |
250 |
68 |
232 |
3 - |
388 |
524 |
331 |
445 |
232 |
602 |
191 |
276 |
205 |
332 |
175 |
269 |
5 - |
61 |
55 |
169 |
126 |
36 |
65 |
28 |
27 |
40 |
31 |
35 |
28 |
Totale |
613 |
881 |
596 |
788 |
366 |
1.046 |
375 |
728 |
296 |
623 |
289 |
547 |
Tabella 3. Distribuzione (N e %) delle allodole nelle diverse classi di grasso, separate per sesso. |
||||||||||||
I dati relativi all’accumulo del grasso sottocutaneo presentano una certa variabilità da un anno all’altro, tuttavia le percentuali relative ad uccelli privi di grasso visibile (classe 0) sono sempre molto basse, mentre in tutti gli anni risulta elevato il numero di uccelli inclusi nelle classi di grasso 3-
Bibliografia
Chiron F. (2002) Variations de la pression de chasse sur un oiseau migrateur en déclin: l’Alouette des champs Alauda arvensis. Rapport DEA Environnement: milieux, techniques, sociétés. 57 p.
Hargues R., Girardot P., Ibañez F., Mourguiart P. and Recarte J. (2007) Migration of the Skylark Alauda arvensis along the southern French Atlantic coast. Wildlife Biology in Practice 3: 93-
Kaiser A. (1993) A new multi-
Petersen B.S. (2007) Management Plan for Skylark
Scebba S. (2001) Biometrics and sex ratios of Skylarks Alauda arvensis during migration in southern Italy. Ringing & Migration 20: 364-
Scebba S. (2009) Rapporto “Alauda 2000”. Indagine sulla migrazione autunnale nella Piana del Volturno (Caserta) in base all’attività di inanellamento: autunno 1998 – 2007.
Scebba S., Soprano M. & Sorrenti M. (2015) Sex-