Calidris ferruginea - Gruppo Inanellamento Limicoli (GIL, Napoli)
Menu principale:
- Home Page
- L'Associazione
-
Ricerche in atto
- Migrazione autunnale Allodola
- Migrazione primaverile Tordo
- Ali Tordo bottaccio in Campania
- Stazione Monte Vaccaro
- Riserva Naturale Foce Volturno
- Inanellamento Medio Volturno
- Avifauna svernante
- Avifauna Foce Neto e Sila
-
Ricerche concluse
- Ali Turdidi e Beccacce ATC LT 2
- Stazione Malga Rosello
- Migrazione Caradriiformi
- Progetto Rondine
- Migrazione autunnale Tordo
- Ali Turdidi e Beccacce ATC CE
- Progetto Alauda 2000
- Osservatorio Roccolo Ganda
- Atlante Migrazioni Puglia
- Download
- News
Calidris ferruginea
Migration pattern and fat reserves of Curlew Sandpipers Calidris ferruginea at a stopover site in southern Italy
Sergio Scebba & Giancarlo Moschetti
International Wader Studies (2006) 19:138-
The Volturno plain, southern Italian coast, is a regular stopover site for Curlew Sandpipers during northward migration. During 1992-
INTRODUCTION
Curlew Sandpipers Calidris ferruginea breed along the high Arctic coastal regions in central and eastern Siberia. The non-
The Volturno plain, a stopover site in the central Mediterranean on the southern Italian coast, is regularly visited by large flocks of Curlew Sandpipers during northward migration, while on southward migration, only small flocks appear. Studies on wader migration in this area have been carried out by the "Gruppo Inanellamento Limicoli (Naples)" since 1991 (Scebba et al. 1994).
The present paper provides information obtained by catching and ringing Curlew Sandpipers on the Volturno Plain during northward migration. We describe a new visual method to score fat deposition in waders, the timing and intensity of migration, morphometric data and details of migration routes, as reflected by recaptures.
STUDY SITE AND METHODS
The Volturno Plain, Caserta (southern Italy; Fig. 1), is an area between Lake Patria and the mouth of the Volturno River which contains many artificial ponds varying in size from 0,5 ha up to c.5 ha. The water supply to these ponds is either from an artesian well or pumped from the nearby watercourse. These ponds were constructed by local hunters for waterfowl shooting from September to March. After this date, the ponds are drained and weed and grass species allowed to grow before being flooded again in September.
Captures occurred twice-
On capture, the birds were placed in cages and later individually marked with alloy rings of the "Istituto Nazionale per la Fauna Selvatica ringing scheme" (Bologna-
The following measurements were taken for each bird: bill length (measured with calipers from bill-
Fat was scored to a simple scale from 0 to 4, based on the observation that in waders there is fat deposition in the area between the articulation of the wing and the articulation of the leg. Fat deposits were estimated visually by blowing the feathers down the wing, and scored as follows: 0 = no fat on the body (body appears blood red); 1 = little yellowish fat visible in the depression up to the articulation of the leg; 2 = as 1 plus fat in a little yellowish triangle between the depression and the articulation of the wing; 3 = fat covered all parts mentioned above, but a small fat-
Because it was not possible to take all measurements on all birds trapped, the total number of measurements does not always equal the number of birds captured. The samples from the seven years were pooled by pentades (Berthold 1973).
For an estimation of potential flight range, four models have been used: the formulae of Mc Neil & Cadieux (1972), Summers & Waltner (1979), Greenewalt (1975) and Davidson (1984), based on estimates of flight speed and metabolism.
RESULTS AND DISCUSSION
Phenology of migration
In total 2,177 Curlew Sandpipers were captured in April and May 1992-
Six of the 2,177 birds were retrapped during southward migration: in Germany (two), Poland (one), Finland (one), Italy (one) and Ukraine (one). During northward migration, three were recaptured in Greece and two in Ukraine. Of these last five birds, two were retrapped after four years, one after two years, one after one year and the last after seven days.
Ringing recoveries occurred along two main routes and confirmed that most of the Curlew Sandpipers staging on the Volturno Plain used other stopover sites in Greece and in the Black Sea area, while on southwardmigration followed the East Atlantic Flyway to their nonbreeding grounds (Fig. 1). The different numbers of birds recorded migrating northward and southward, confirms this migration strategy along two different routes.
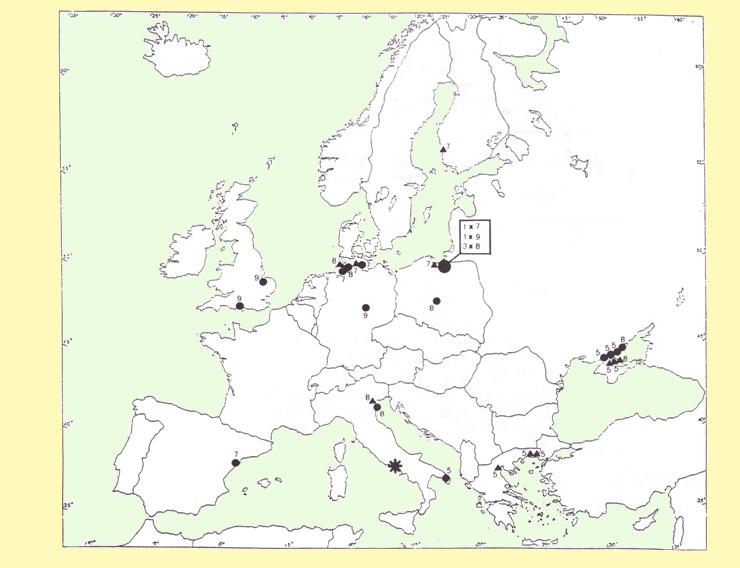
Figure 1. Ringing and recovery sites of Curlew Sandpipers recovered (circles) or ringed (triangles) in spring in Volturno plain. Numbers denote ringing month (for foreign birds) and recovery month (for birds ringed in Volturno plain). Asterisk= study area.
All the birds were caught between 16 April to 30 May (pentades 22-
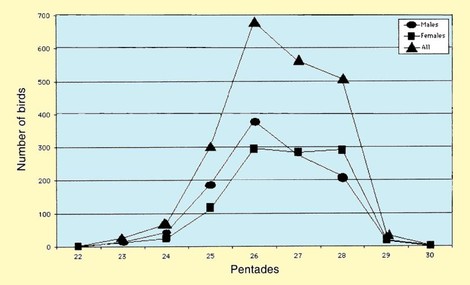
Figure 2. Trapping patterns, per pentade, of Curlew Sandpipers on the Volturno plain during northward migration 1992-
The numbers of Curlew Sandpipers caught over the years fluctuated markedly (Table 1); similar trends were found in Tunisia during the period 1966-
The median dates of passage for males and females were in pentades 26 (6-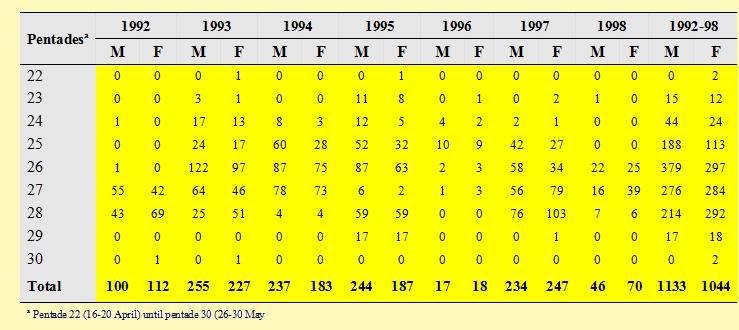
Table 1. Numbers of Curlew Sandpipers trapped on the Volturno plain per year and per pentades during spring 1992-
Site fidelity
It is possible that the same flocks visited this stopover site annually. We recaptured, at the same site, seven birds ringed during previous northward migrations. Particularly, two birds ringed on 30 April 1993 and 1 May 1993, were recaptured after one year on the same date. Other birds, that were first ringed on the same day in Poland (Vistula mouth, near Gdansk), were also recovered on the Volturno Plain.
Sex ratios during migration
In order to confirm the effectiveness of the Wymenga et al.'s (1990) discriminant function, 100 individuals were sexed only on the basis of plumage characteristics: female have pale pink underparts, while males have dark vinaceous-
Age structure during migration
All trapped birds were aged on the basis of plumage characteristics (Prater et al. 1977); only 20 birds (0.9%) were second-
Biometric data
The monotypic Curlew Sandpiper does not show any geographical variation in size. Table 2 summarizes measurements of wing, third primary, tarsus, bill, nalospi, total head and body mass of Curlew Sandpipers captured on the Volturno Plain during 1992-
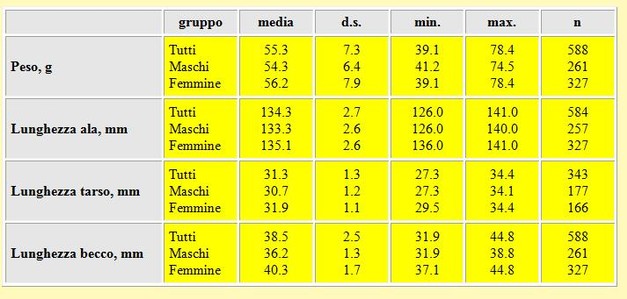
Table 2. Summarized data of measurements (in mm) and body mass (g) of Curlew Sandpipers trapped on the Volturno Plain 1992-
The difference in wing length between males and females was highly significant (t = 31.0, P < 0.01), as were differences in third primary, tarsus, bill, nalospi and total head length (t-
The mean of wing length, on the Volturno Plain did not differ from that in the Sivash, Ukraine (mean 133,3 mm, SD 3.1, n = 224; Chernichko et al. 1993), but differed by 0.6 mm from wing lengths presented in Meininger et al. (1990) for northeastern Greece (mean 132.7 mm, SD 2.8, n = 85). The mean total head length did not differ significantly from the measurements collected in Ukraine (mean 61.5 mm, SD 2.72, n = 26; Chernichko et al. 1993) (t =1.6, n.s.) and in northeastern Greece (mean 62.1 mm, SD 2.64, n = 87; Meininger 1990) (t = 0.4, n.s.). The mean bill length did not differ significantly from the measurements collected in northeastern Greece (mean 38.1 mm, SD 2.41, n = 87, Meininger 1990), but was significantly smaller than measured in the Ukraine (mean 39.0 mm, SD = 3.0, n = 224; Chernichko et al. 1993) (t = 8.3, P < 0.01). The data collected on the Volturno Plain for males and females were not different from data presented by Cramp & Simmons (1983). Tarsus length presented in this study tended to be greater than that reported in Cramp & Simmons (1983). Fig. 3 shows frequency distributions of the wing (Fig. 3A), third primary (Fig. 3B), tarsus lengths (Fig. 3C) and bill (Fig. 3D), in which sexed birds are indicated separately. A clear bimodality becomes apparent, especially in bill length. This bimodality results from sexual dimorphism. 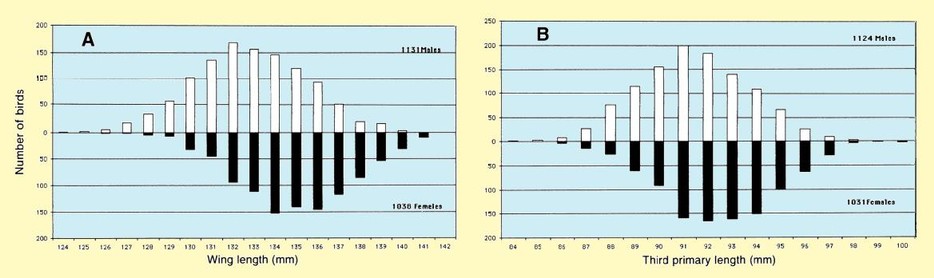
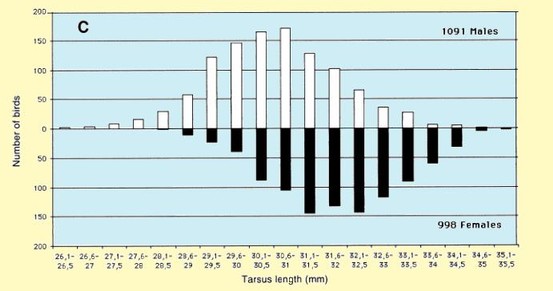
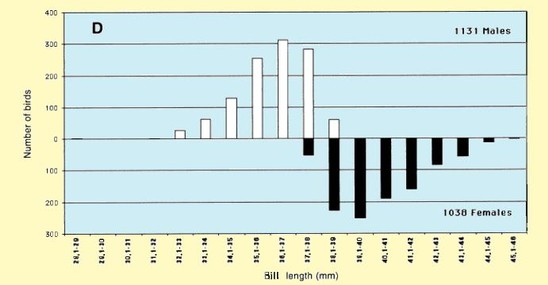
Figure 3. Frequency distributions of (A) wing length (in 1-
Analysis of body mass
The mean body mass of 54.3 g (Table 2) on the Volturno Plain was considerably lower than in Camargue, France in May (mean 60.8 g, SD 8.3, n = 66; Glutz et al. 1975) and in Sivash, Ukraine (mean 56.3 g, SD 7.0, n = 221; Chernichko et al. 1993). These values are statistically different from ours (t-
Variation in body mass per pentade is shown in Fig. 4. Only four years with relatively large numbers of captures are considered. In these four years, there is a large fluctuation in weights per pentade. This can be attributed to continuous immigration of new birds from Africa. The means of weights calculated for males per pentade (Fig. 4A), showed a similar pattern to that of females (Fig. 4B). The mean body mass of males was 53.5 g (SD 6.9, n = 1128, min. 36.0 g, max 81.5 g), while the mean for females was 55.1 g (SD 7.0, n = 1030, min 39.0 g, max 82.3 g). The difference between the means was statistically significant (t-
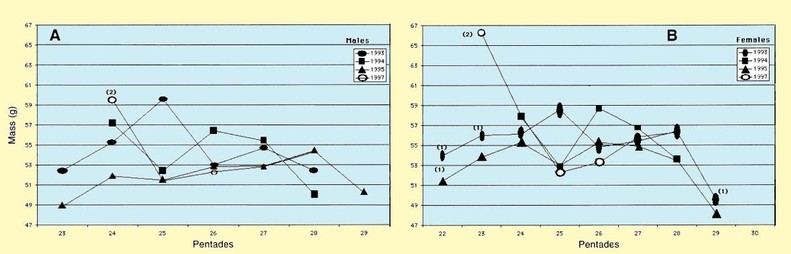
Figure 4. Variation in body mass, per pentade, of Curlew Sandpipers trapped during northward migration 1993, 1994, 1995 and 1997. Pentades 22 = 16-
Fat score
In this study we attempted to carry out a new visual method to evaluate the fat deposition in some wader species. Therefore, as reported under Methods, we attributed a fat score on the basis of fat accumulation. The results of the corresponding mean weights per fat score are presented in Table 3.
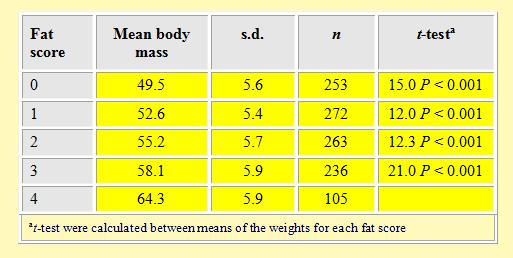
Table 3. Mean weight for each fat score of Curlew Sandpipers trapped at Volturno plain between 1993-
There were a significant relationships between the body weight of birds and visually estimated fat deposits: higher weights corresponded to a higher fat scores, with a statistically significant difference between the mean weights per fat score. Moreover, there was a strong correlation between fat scores and mean weight per fat score (Pearson coefficient = 0.982).
Body mass variation of retrapped birds and length of stay
Only 34 birds out of 2,177 were caught twice within the same period. There was a substantial change in body mass: 22 birds recovered after 0.5-
On the basis of the days that elapsed between ringing and recapture, mean stopover duration was estimated to be 2.2 days for pre-
Flight range
A considerable proportion of the Curlew Sandpipers migrating through the Volturno Plain use wetlands in Greece and southern Ukraine as stopover areas before flying to the breeding grounds (Fig. 1). In order to confirm our assumption, the potential flight range of Curlew Sandpipers leaving the Volturno Plain was evaluated.
For an estimation of potential flight range, both mass at arrival (lean mass) and departure mass (total mass at start) must be known. Many authors (Zwarts et al. 1990, Meininger 1994) used a lean mass calculated as the average mass recorded at nonbreeding areas in the tropics, and departure mass calculated as the mean of the upper half of the frequency distribution of weight of the birds "ready to go" (birds that have finished moult). In our study area, Curlew Sandpipers showed a varying mixture of old and new feathers without active moult of body or tail feathers; in addition, the length of stay on the Volturno Plain was short. As a consequence, there was no sound basis for identifying birds that were "ready to depart". Therefore, we used as lean mass and as departure mass the mean weight calculated respectively for the lowest and for the highest class of fat score (Table 3).
At the flight speed predicted by Pennycuick (1989), 41.2 km/h, the four models used yielded still-
However, the distances mentioned above could be under-
CONCLUSIONS
The following points emerge from the data on Curlew Sandpipers ringed on the Volturno Plain:
1) The ringing area represents an important stopover site for wader species (Scebba et al. 1994, Scebba & Moschetti 1996), especially for Curlew Sandpiper during northward migration. The level of brackish water in artificial ponds is sufficiently constant and the quality of water causes large numbers of crustaceans (Gammarus sp.) and of aquatic dipteran larvae (Chironomidae) to be present. When the ponds are drained, these arthropods aggregate in the remaining shallow puddles, creating a concentratedsource of food. This occurrence coincides with northward migration.
2) During northward migration, large flocks of Curlew Sandpipers stop on the Volturno Plain, but only sporadically on southward migration. In early May, it is not uncommun to see flocks of 2,000-
3) Analysing the map of bird recoveries during southward migration, it seems that Curlew Sandpiper follow the East Atlantic Flyway along the Baltic coast to their nonbreeding grounds. During northward migration, these birds use the central Mediterranean Flyway, using wetlands in southern Italy, Greece and the Black Sea, before arriving at the breeding grounds.
4) Curlew Sandpipers showed a mean mass increase of 2.8 g/day and the mean stopover duration was 2.2 days at the study site.
5) It is possible that the same flocks visit in spring this stopover site annually, because we controlled four birds ringed during the previous pre-
6) Only 0.9% of the total catch of Curlew Sandpipers were first-
7) A flight range of between 1,000 and 1,900 km would allow Curlew Sandpipers to leave the Volturno Plain, cross the Adriatic Sea and reach the wetlands along the southern coast of Greece (c. 1,000 km), the Danube Delta (c. 1,300 km) and northern Black Sea coast (c. 1,700 km).
ACKNOWLEDGEMENTS
Rodney West, Paul Newton, Michael Wright, John Glazebrook and Alan Miller of the InternationalWader Study Group, Phil Ireland of the Wash Wader Ringing Group, Zsolt Karcza and Tibor Csorgo of the Hungarian Ornithological Society, Wlodzimierz Meissner and Mateusz Sciborski of the "Grupa Badawcza Plakow Wodnich KULING", Poland, Stefano Laurenti of Società Ornitologica Italiana (SOI), Roy Hughes, Jan Spence and Gianluigi Mauriello helped in the field. Gabor Lovei reviewed the text and Mark Walters made stylistic amendments. Daniele Anesa and the "Roccolo Ganda" Bird Observatory (Aviatico-
REFERENCES
Berthold, P. 1973. Proposals for the standardization of the presentation of annual events, especially of migrating data. Auspicium, 5 (suppl.): 49-
Chernichko, I.I., Chernichko, R., Diadicheva, E., van der Have, T.M. & van de Sant, S., 1993. Biometry of waders in the Sivash, Ukraine, spring 1992. In: van der Have, T.M., van de Sant, S., Verkuil, Y. & van der Winden, J. (Eds.) Waterbirds in the Sivash, Ukraine, spring 1992. WIWO-
Cramp, S. & Simmons, K.E.L. 1983. The Handbook of the birds of the western Palearctic. Vol. 3. Oxford: Oxford University Press.
Davidson, N.C. 1984. How valid are flight range estimates for waders? Ring. & Migr. 5: 49-
Glutz von Blotzheim, U.N., Bauer, K.M. & Bezzel, E. (Eds). 1975. Handbuch der Voegel Mitteleuropas.
Greenewalt, C.H. 1975. The flight of birds. Trans. Amer. Phil. Soc. 65: 1-
Mc Neil, R. & Cadieux, F. 1972. Numerical formulae to estimate flight range of some American shorebirds from fresh weight and wing length. Bird-
Meininger, P.L. & Schekkerman, H. 1994. Measurements, moult and mass of waders captured in Egypt, winter and spring 1990. In: Meininger P.L. & Atta G.A.M. (Eds.). Ornithological studies in Egyptian wetlands 1989/90. WIWO Report
Meininger, P.L., Spiekman, H.W. & van Westrienen, R. 1990. Measurements and plumage of waders captured in NE-
Norrevang, A. 1959. The migration patterns of some waders in Europe, based on the ringing results. Vidensk. Medd. Dansk. Naturh. Foren.
Pennycuick, C.J. 1989. Bird Flight Performance: a Practical Calculation Manual.
Prater, A.J., Marchant, J.H. & Vuorinen, J. (1977) Guide to Identification and Ageing of Holarctic Waders. B.T.O. Guide 17. Tring: British Trust for Ornithology.
Scebba, S. 1993. Gli Uccelli della Campania. Napoli: Ed. Esselibri.
Scebba, S., Moschetti, G. & Taglialatela, D. 1994. Importanza delle vasche artificiali nella Piana del Volturno (Caserta) per la migrazione dei Caradriformi. In: Fraissinet M., Coppola D., Del Gaizo S., Grotta M. e Mastronardi D. (Eds) Atti Convegno Nazionale "L'avifauna degli ecosistemi di origine antropica: zone umide artificiali, coltivi, aree urbane". Monografia 5 ASOIM, Electa Napoli: 151-
Scebba, S. & Moschetti, G. 1996. Migration pattern and weight change of Wood Sandpiper Tringa glareola
Spiekman, H.W. 1992. Results of Wader Ringing Activities in Tunisia 1962-
Summers, R.W. & Waltner, M. 1979. Seasonal variations in the mass of waders in southern Africa, with special reference to migration. Ostrich 50: 21-
Wylson, J.R., Czajkowski, M.A., Pienkowski, M.W. 1980. The migration through Europe and wintering in West Africa of the Curlew Sandpiper. Wildfowl
Wymenga, E., Engelmoer, M., Smit, C.J. & van Spanje, T.M. 1990. Geographical breeding origin and migration of waders wintering in West Africa. Ardea
Zwarts, L., Ens, B.J., Kersten, M. & Piersma, T. 1990. Moult, mass and flight range of waders ready to take off for long-
Paper n.29 of Gruppo Inanellamento Limicoli (G.I.L., Naples), Italy.