Resoconto 2002 - Gruppo Inanellamento Limicoli (GIL, Napoli)
Menu principale:
- Home Page
- L'Associazione
-
Ricerche in atto
- Migrazione autunnale Allodola
- Migrazione primaverile Tordo
- Ali Tordo bottaccio in Campania
- Stazione Monte Vaccaro
- Riserva Naturale Foce Volturno
- Inanellamento Medio Volturno
- Avifauna svernante
- Avifauna Foce Neto e Sila
-
Ricerche concluse
- Ali Turdidi e Beccacce ATC LT 2
- Stazione Malga Rosello
- Migrazione Caradriiformi
- Progetto Rondine
- Migrazione autunnale Tordo
- Ali Turdidi e Beccacce ATC CE
- Progetto Alauda 2000
- Osservatorio Roccolo Ganda
- Atlante Migrazioni Puglia
- Download
- News
Resoconto 2002
Monitoraggio delle popolazioni di limicoli nella Piana del Volturno (Caserta) durante la migrazione primaverile del 2002
Sergio Scebba
Gli Uccelli d'Italia XXVIII (1-
Introduzione
Il termine "flyway" viene impiegato per individuare determinate aree del mondo utilizzate dai Caradriformi e comprende sia le rotte migratorie che le aree di sosta usate dalle diverse popolazioni nel corso dei loro movimenti tra le zone di nidificazione e quelle di svernamento. Ciascuna popolazione migra secondo un suo caratteristico schema ed utilizza un particolare insieme di punti di riproduzione, di sosta e di svernamento. Per tale motivo una flyway risulta composta da tanti sistemi di migrazione quante sono le popolazioni o le specie di limicoli che la percorrono, secondo le rispettive preferenze ambientali e secondo le strategie migratorie adottate (Hötker et al. 1998).
L'Italia, per la sua posizione geografica, rappresenta un ponte ideale verso il continente africano e rientra a far parte della "Mediterranean/Black Sea Flyway". Gli uccelli che utilizzano gli habitats mediterranei sia per brevi soste, onde ripristinare le riserve energetiche necessarie, che per l'intero periodo dello svernamento, nidificano nella regione artica e subartica dell'Europa settentrionale e della Siberia, nella regione temperata ed anche lungo le coste del Mar Nero e del Mediterraneo. A causa della scomparsa lungo il tratto sud-
Un notevole interesse è attualmente rivestito da numerosi invasi artificiali localizzati nella Piana del Volturno in provincia di Caserta. In quest'area il Gruppo Inanellamento Limicoli (G.I.L., Napoli), con la collaborazione del Wader Study Group, dell'Hungarian Ornithological Society e del Kuling Group (Polonia), ha avviato dal 1991 un programma di studio a lungo termine (Scebba et al.
I dati raccolti sono di grande rilevanza scientifica in quanto in Italia meridionale, ed in Campania in particolare, i movimenti migratori dei Caradriformi non sono mai stati sufficientemente indagati ed anche l'inanellamento è sempre avvenuto in modo occasionale. Le informazioni relative alle disponibilità alimentari ed alle variazioni di peso nelle aree di sosta durante le migrazioni sono praticamente inesistenti ed anche le conoscenze sulle popolazioni di limicoli e sul numero di individui coinvolti sono ancora molto incomplete. La ricerca si propone quindi di:
-
-
-
-
-
In questo studio viene presentata un'analisi preliminare dei dati raccolti nel corso della migrazione primaverile del 2002.
Materiali e metodi
La ricerca è stata condotta in un'area costituita da due località denominate "Soglitelle" e "Pericarole" comprese nel comune di Villa Literno (Tavoletta I.G.M. 184 IV NO; coordinate 40.58N 13.59E; superficie c/a 190 ha). In quest'area è presente un elevato numero di invasi artificiali, utilizzati in gran parte per scopi venatori da Settembre a Gennaio, di dimensioni e profondità molto variabili (da qualche centinaio di mq fino a 4-
Il progetto è stato articolato su due linee principali di ricerca:
1) attività di cattura ed inanellamento;
2) osservazioni.
L'attività di cattura ed inanellamento è stata effettuata dal 29 Marzo al 26 Aprile e dal 10 al 18 Maggio con una frequenza di due giorni per pentade, in quanto i tassi di cattura tendono a decrescere notevolmente dopo il secondo giorno di attività (Karr 1981); dal 27 Aprile al 9 Maggio, periodo di maggiore intensità migratoria caratterizzato da un considerevole turn-
Le catture sono avvenute di notte con reti mist-
Alla cattura, gli uccelli sono stati posti in sacchetti di tela o in gabbie completamente ricoperte e quindi subito inanellati con anelli dell'Istituto Nazionale per la Fauna Selvatica (Bologna-
Su tutti gli individui catturati sono state rilevate di routine alcune misure biometriche: lunghezza del becco (misurata con un calibro dalla punta del becco alle prime piume, Svensson 1992) con approssimazione di 0,1 mm; nalospi (misurata con un calibro dalla punta del becco alla narice, Prater et al. 1977) con approssimazione di 0,1 mm; lunghezza dell'ala (misurata secondo il metodo della corda massima, appiattita e raddrizzata, Svensson 1992) con approssimazione di 0,5 mm; lunghezza della III remigante primaria (massima lunghezza, Berthold & Friedrich 1979) con approssimazione di 0,5 mm; lunghezza del tarso (misurata con un calibro, Svensson 1992) con approssimazione di 0,1 mm; peso (a mezzo di bilancina elettronica) con approssimazione di 0,1 gr.
L'età è stata determinata in base alle caratteristiche del piumaggio, al consumo delle remiganti primarie ed al tipo di muta effettuata (Prater et al. 1977). La stima della quantità di grasso visibile è stata stabilita secondo una scala da 0 a 4, in base alla quantità presente tra l'articolazione dell'ala e quella della zampa (Moschetti & Scebba 1999, Scebba & Moschetti in stampa). La muta delle penne del corpo è stata registrata usando una scala da 0 a 2 (0, nessuna penna in crescita; 1, fino a 20 penne in crescita; 2, più di 20 penne in crescita). Tutta l'operazione non ha richiesto più di un'ora, tempo massimo entro il quale tutti gli uccelli catturati sono stati di nuovo liberati.
I dati raccolti sono stati raggruppati e distribuiti in periodi di 5 giorni (pentadi) (Berthold 1973). Allo scopo di stimare l'autonomia di volo potenziale degli uccelli che sostano nell'area è stato utilizzato un modello matematico (Davidson 1984) basato sulla velocità di volo, sul peso all'arrivo (peso magro) e sul peso alla partenza (massa totale degli uccelli pronti a migrare).
La formula è:
R = 95.447 * S * (T 0.302 -
in cui S = velocità di volo (km/h), T = peso alla partenza, M = peso magro.
Molti autori (Zwarts et al.
Le osservazioni sono state condotte quotidianamente ed effettuate sia a piedi che in auto; i conteggi sono avvenuti al tramonto, quando gli uccelli erano intenti ad alimentarsi o a riposare. Sono stati utilizzati binocoli 8x40 e 10x40, cannocchiale 20-
Sono stati ancora rilevati i dati climatici, quali pressione atmosferica, umidità relativa, temperatura, copertura del cielo, piovosità, intensità del vento. Tutti i dati raccolti sono stati quindi riportati su schede prestampate e successivamente archiviati in un Data-
Risultati
Analisi dei dati
Nel corso della prima stagione del progetto sono stati inanellati 1.559 uccelli appartenenti a 27 specie. Il Piovanello (Calidris ferruginea) ha caratterizzato il 2002, in quanto ha costituito il 37% dell'intera popolazione inanellata. È importante rimarcare la cattura di alcune specie di notevole importanza per l'intero territorio nazionale e regionale, incluse nell'Allegato I della Direttiva 79/409/CEE ed in particolare il Piro piro boschereccio (Tringa glareola), il Cavaliere d'Italia (Himantopus himantopus), il Combattente (Philomachus pugnax), il Mignattino (Chlidonias niger) ed il Tarabusino (Ixobrychus minutus); per quest'ultima specie, due degli individui catturati presentavano la placca incubatrice, segno della loro nidificazione nei radi canneti che fiancheggiano i numerosi canali della zona. Sono state anche inanellate alcune specie rare o comunque poco comuni quali il Gambecchio frullino (Limicola falcinellus), l'Albastrello (Tringa stagnatilis) ed il Voltapietre (Arenaria interpres). L'elenco completo degli inanellamenti è presentato in tabella 1.
Tabella 1. Elenco delle specie inanellate nel periodo 29 Marzo/18 Maggio 2002.
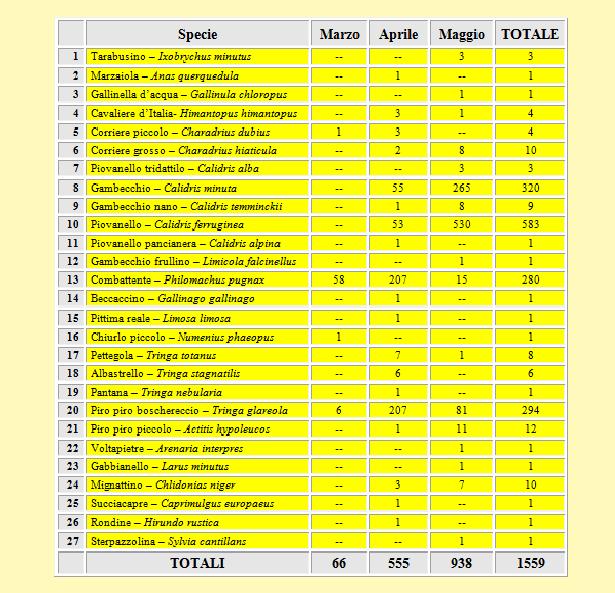
L'intera stagione di studio si è realizzata in 29 uscite in campo effettive, che hanno prodotto una quota media di 54 uccelli per ciascuna uscita con un valore minimo di un solo inanellato ed un valore massimo di 193 catture. Sono stati inanellati uccelli appartenenti a 25 specie di non Passeriformi (di cui 21 Caradriformi) ed a 2 specie di Passeriformi. Una rappresentazione grafica delle specie maggiormente inanellate è mostrata in Figura 1.
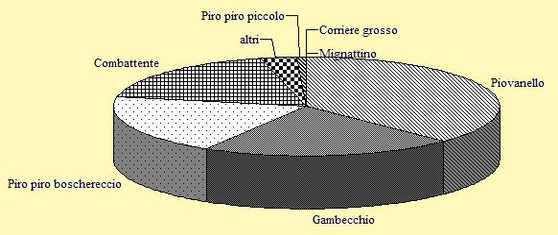
Figura 1. Percentuale delle specie inanellate nel periodo 29 Marzo/18 Maggio 2002 (n= 1560).
In totale, dal 29 Marzo al 18 Maggio, sono stati catturati 1.560 uccelli appartenenti all'ordine dei Caradriformi (questo numero comprende anche le ricatture di quattro uccelli con anello estero e di cinque individui inanellati nel corso degli anni precedenti). La data mediana delle catture (quando cioè è stato catturato il 50% di tutti gli uccelli) si è verificata tra il 2 ed il 3 Maggio (pentade 25); il primo quartile, che corrisponde al 25% delle catture, è terminato tra il 21 ed il 23 Aprile (pentade 23) ed il terzo quartile ha avuto inizio tra il 10 e l'11 Maggio (pentadi 26-
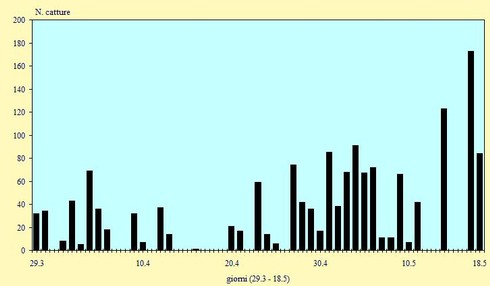
Figura 2. Andamento % delle catture per pentade nel periodo 29 Marzo/18 Maggio 2002 (n=1560).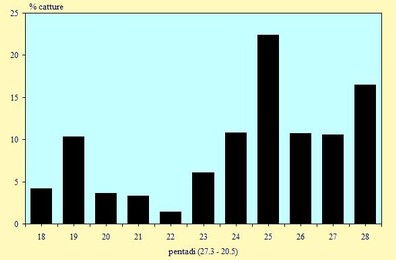
Figura 3. Andamento giornaliero delle catture nel periodo 29 Marzo/18 Maggio 2002 (n=1560).
Nel periodo di studio la migrazione è stata caratterizzata dalla presenza di tre diverse ondate: la prima ad inizio Aprile (pentade 20), determinata per lo più dal passaggio di Combattenti, soprattutto femmine, in quanto i maschi migrano molto più presto; quindi i numeri sono andati diminuendo (un solo inanellato il giorno 16) per poi risalire nuovamente verso la fine di Aprile (pentade 23) quando le catture sono aumentate con un andamento abbastanza regolare fino ad un forte passaggio agli inizi di Maggio (pentade 25). Una terza ondata migratoria, dovuta ad un intenso transito di Piovanelli e Gambecchi, si è verificata nella seconda metà di Maggio: nella notte tra il 17 ed il 18 sono stati inanellati 193 uccelli.
In base all'esame dei giorni intercorsi tra inanellamento e ricattura di alcuni Gambecchi e Piovanelli, sono stati valutati i tempi medi di sosta nell'area di studio; inoltre, attraverso le ricatture di uccelli inanellati in anni precedenti, è stata confermata la fedeltà a quest'area di sosta dimostrata da migratori su lunga distanza, dove per lunga distanza s'intende il percorso tra l'estrema tundra siberiana, oltre il Circolo Polare Artico, e le coste africane dell'Oceano Atlantico, cioè circa 6000/7000 Km.
Nella figura 4 sono riportate in percentuale i numeri dei Caradriformi osservati (non sono inclusi i Laridi in quanto catturati solo occasionalmente).
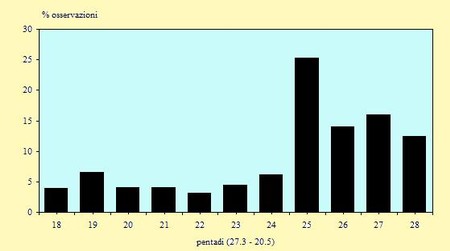
Figura 4. Andamento % delle osservazioni per pentade nel periodo 29 Marzo/18 Maggio 2002.
E' possibile evidenziare che intorno alla metà di Aprile la migrazione, anche se ormai in atto, non presenta ancora numeri molto elevati; il picco migratorio si ha nella prima decade di Maggio, in particolare per l'arrivo di stormi molto numerosi di Gambecchi e Piovanelli. I due grafici, relativi a catture ed osservazioni (Fig. 2 e 4), presentano un andamento abbastanza sovrapponibile; una leggera difformità si nota nelle ultime pentadi, dovuta ad un maggiore impegno nelle operazioni di cattura.
Numerose sono state le osservazioni di specie incluse nell'Allegato I della Direttiva 79/409/CEE: Spatola (Platalea leucorodia), Airone rosso (Ardea purpurea), Pernice di mare (Glareola pratincola), Avocetta (Recurvirostra avosetta), Gabbiano roseo (Larus genei), Sterna maggiore (Sterna caspia), Beccapesci (Sterna sandvicensis), Sterna comune (Sterna hirundo) e Mignattino piombato (Chlidonias hybridus), per le quali sono stati accertati regolari movimenti migratori. È inoltre interessante segnalare il passaggio a fine Marzo di uno stormo di circa un centinaio di Mignattai (Plegadis falcinellus), la sosta dagli inizi di Maggio per almeno due settimane di un gruppo di 32 Fenicotteri (Phoenicopterus roseus), la costante presenza (fino a 150 individui) di numerose Garzette (Egretta garzetta). Per due specie è stata confermata la regolare nidificazione: da diversi anni si è ormai insediata nell'area una colonia (non meno di 90/100 individui) di Cavalieri d'Italia (Himantopus himantopus) e sono regolarmente presenti almeno 5/6 coppie di Pernici di mare (Scebba e Moschetti 1995).
Analisi della migrazione del Gambecchio, Piovanello e Piro piro boschereccio
La fenologia della migrazione espressa in pentadi delle tre specie più inanellate -
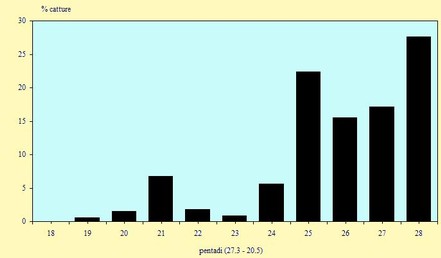
Figura 5. Andamento delle catture, nel periodo 29 Marzo -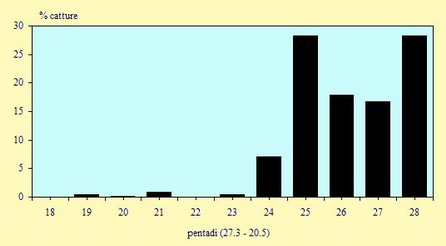
Figura 6. Andamento delle catture, nel periodo 29 Marzo -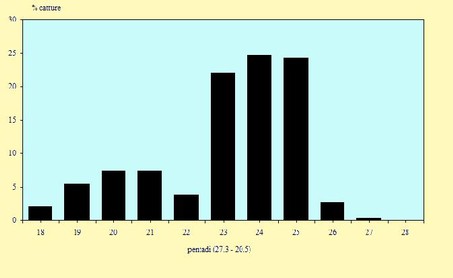
Figura 7. Andamento delle catture, nel periodo 29 Marzo -
La Piana del Volturno rappresenta un'importante zona di sosta lungo la Mediterranean/Black Sea Flyway, in quanto durante la migrazione pre-
Gambecchio Calidris minuta
Questa specie nidifica nella tundra artica, dalla Norvegia settentrionale alla Siberia centrale (Cramp & Simmons 1983). La migrazione avviene su di un largo fronte; in base ai dati forniti dalle ricatture sembrerebbe che le popolazioni che svernano nell'area del Senegal/Gambia durante il ritorno alle aree di riproduzione seguano una rotta che attraversa la Tunisia, il Mediterraneo centrale e l'area del Mar Nero (Wymenga et al. 1990).
Fenologia della migrazione
Le catture sono avvenute tra il 4 Aprile ed il 18 Maggio (pentadi 19-
Il numero degli individui catturati nel corso di questo primo anno non è stato particolarmente elevato, soprattutto se confrontato con quelli ottenuti in alcuni anni precedenti (in particolare nel 1994 -
È stata effettuata una ricattura straniera: un Gambecchio inanellato in Polonia è stato ricatturato il 23 Aprile; inoltre, un uccello inanellato il 15 Maggio 2001 è stato ricatturato ancora a distanza di un anno (1 Maggio 2002), confermando l'importanza dell'area come zona di sosta per questa specie, in particolare durante la migrazione primaverile. Analoghi risultati sono emersi anche per il Piovanello.
Analisi biometrica
Il Gambecchio è una specie monotipica che mostra modeste variazioni nelle dimensioni; pertanto attraverso i dati morfometrici è molto difficile determinare l'origine geografica delle diverse popolazioni. In tabella 2 sono raccolte le misure relative a peso, lunghezza alare, tarso e becco degli uccelli catturati.
Tabella 2. Peso (in g) e misure biometriche (in mm) dei Gambecchi, primavera 2002.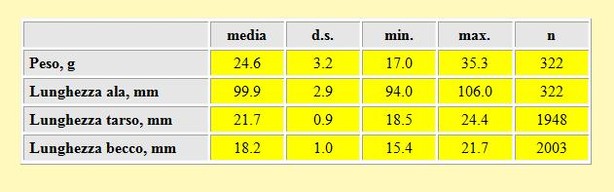
La lunghezza alare media calcolata per tutti gli uccelli è stata di 99.9 mm (d.s. 2.9, min 94mm, max 106 mm, n= 322). Cramp & Simmons (1983) riportano una distribuzione più ridotta di 93-
Il peso medio calcolato per l'intero periodo di cattura (4 Aprile-
Si è quindi tentato di valutare se erano presenti delle differenze di dimensione tra gli uccelli catturati nel primo periodo della migrazione (pentadi 19-
Tabella 3. Confronto tra le medie delle lunghezze dell'ala (mm) e del becco (mm) dei Gambecchi, primavera 2002, durante l'inizio (pentadi 19-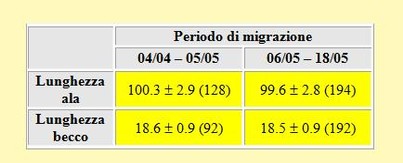
Dalla tabella si evidenzia una chiara presenza di uccelli con ali più lunghe nella prima parte della migrazione rispetto a quelli catturati nel secondo periodo, cioè dopo il 5 Maggio. Le differenze tra le lunghezze alari calcolate nei due periodi sono state altamente significative. Al contrario, non sono state trovate differenze per quanto riguarda le medie della lunghezza del becco calcolate prima e dopo il 5 Maggio. I nostri risultati sono discordanti rispetto a quanto riportato da Chernichko et al. (1993) per il Sivash. In effetti questi autori hanno trovato nella seconda metà di Maggio uccelli con l'ala più lunga ma con becco più corto. Purtroppo non è chiaro se queste differenze vanno imputate a differenze tra sessi oppure, più probabilmente, a differenze geografiche dovute a popolazioni più orientali che passano prima.
Sesso ed età
Non esistono differenze tra i sessi per quanto riguarda il piumaggio, ma vi sono delle modeste differenze nelle dimensioni, in quanto le femmine si presentano leggermente più grandi; in ogni caso, in campo non è possibile stabilire il sesso (Wymenga et al. 1990, Meininger & Schekkerman 1994). Una parte dei Gambecchi catturata può però essere sessata utilizzando la lunghezza del becco e dell'ala (maschi: becco < 17 mm, ala < 95 mm; femmine: becco > 19 mm, ala > 99 mm). Usando questi criteri 80 (28.1%) su 285 Gambecchi sono stati identificati come femmine e solo 2 (0.7%) come maschi. Analoghi risultati (23.6% e 0.5%) sono stati accertati per la Grecia nord-
Analisi del grasso
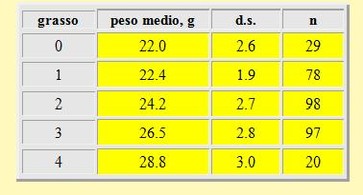
I risultati dei corrispondenti pesi medi per ciascuna classe di grasso sono presentati nella tabella 4. Vi è una significante relazione tra il peso medio e la quantità di grasso stimata visualmente: le medie più alte corrispondono evidentemente alle classi di grasso maggiori, con differenze statisticamente significanti tra le medie di ciascun peso per classe di grasso (t-
Tabella 4. Peso medio per ciascuna classe di grasso nei Gambecchi, primavera 2002.
Variazione del peso nei ricatturati e lunghezza del periodo di sosta
Nel corso delle operazioni di cattura sono stati ricontrollati soltanto due individui su 322 catturati, a distanza di tre giorni dall'inanellamento. I due uccelli hanno guadagnato peso, mostrando un incremento di 0.9 g/giorno, un valore molto alto se confrontato con quelli riportati in letteratura per aree dove avviene l'accumulo di grasso premigratorio (Meininger & Schekkerman 1994).
Sulla base dei giorni intercorsi tra cattura e ricattura è evidente che il periodo medio di sosta nell'area è molto breve, ma rispecchia comunque quanto già accertato in altre aree, quali Grecia e Turchia. Analoghi risultati sono stati ottenuti per il Piovanello.
Autonomia di volo
Utilizzando i seguenti valori di peso è stato calcolata l'autonomia di volo potenziale:
Peso magro (senza accumulo di grasso) = 22.0 g;
Peso alla partenza (con accumulo di grasso) = 28.8 g.
Zwarts et al. (1990) e Meininger (1994) hanno utilizzato per il peso magro il peso medio rilevato nelle aree tropicali di svernamento (22.1 g), valore molto simile a quello calcolato in questa ricerca; diversamente, il peso degli uccelli pronti a migrare accertato in Egitto (Meininger 1994) risulta molto superiore al nostro. Con il modello matematico utilizzato (Davidson 1984) e con una velocità di volo di 41.2 km/h ipotizzata da Pennycuick (1989) è stato quindi possibile stimare un'autonomia di volo potenziale di 845 km per i Gambecchi che lasciano la Piana del Volturno. Ad una velocità di volo di circa 55 km/h accertata per i Limicoli e basata su osservazioni in campo (Zwarts et al. 1990) tale autonomia può arrivare a 1129 km.
Naturalmente queste distanze sono più ridotte rispetto a quelle ipotizzate per gli uccelli che lasciano l'Egitto. Qui è necessario fare due considerazioni. La prima è che una volta raggiunta l'Italia meridionale gli uccelli possono adottare una strategia di volo che prevede tragitti di media lunghezza considerato che, superato l'ostacolo del Sahara e del Mediterraneo, possono trovare comunque delle aree umide in cui sostare. La seconda considerazione è che mentre la classe di grasso 0 non presenta quasi per niente grasso (o comunque in quantità insufficiente per la migrazione), la classe 4 potrebbe includere individui con quantità di grasso tali da rendere indispensabile l'introduzione di un'ulteriore classe 5; di conseguenza il peso medio alla partenza da noi utilizzato potrebbe essere sottostimato. Comunque un'autonomia di volo potenziale variabile tra 800 e 1100 km consentirebbe ai Gambecchi che lasciano la Piana del Volturno di attraversare l'Adriatico per raggiungere altre aree umide quali, ad esempio, quelle localizzate tra il delta del Nestos ed il delta dell'Evros lungo le coste nord-
Piovanello Calidris ferruginea
Questa specie nidifica lungo le regioni costiere del Circolo Polare Artico della Siberia centrale ed orientale; l'areale di svernamento va dall'Africa occidentale all'Australia, ma durante l'inverno le maggiori concentrazioni sono presenti nella fascia costiera dell'Africa equatoriale (Cramp & Simmons 1983). Ricatture di uccelli inanellati vengono segnalate lungo due rotte principali: lungo la fascia costiera dell'Europa occidentale e più internamente attraverso la Tunisia ed il Mar Nero; una parte della popolazione che nidifica in Siberia segue questa rotta più orientale, conosciuta come Mediterranean/Black Sea Flyway (Wilson et al. 1980).
È stato accertato che in primavera, per ritornare nei territori di riproduzione, gli uccelli che hanno svernato nell'Africa occidentale sorvolano il Sahara ed attraversano il bacino del Mediterraneo utilizzando lungo questa rotta una serie di aree di sosta localizzate in Tunisia, in Italia meridionale, in Grecia e nell'area del Mar Nero.
Fenologia della migrazione
Le catture sono avvenute tra il 2 Aprile ed il 18 Maggio (pentadi 19-
Il numero dei Piovanelli inanellati nel corso di questo primo anno del progetto è risultato il più alto rispetto a quelli realizzati negli anni precedenti. Queste fluttuazioni rientrano comunque nel normale andamento della fenologia della migrazione in quanto, da un anno all'altro, è possibile che si verifichino delle grosse variazioni nei passaggi dei migratori.
Come già è stato evidenziato dai dati raccolti negli anni precedenti (Moschetti e Scebba 1999), i maschi hanno mostrato la tendenza a precedere le femmine: durante il periodo principale della migrazione (26 Aprile/20 Maggio, pentadi 24-
È stata effettuata una ricattura straniera: un uccello inanellato dal centro d'inanellamento di Bruxelles (Belgio) è stato ricatturato il 5 Maggio. È possibile che gli stessi gruppi tendono a ritornare ogni primavera in quest'area di sosta in quanto, come già avvenuto diverse volte in passato, sono stati ricontrollati quattro uccelli inanellati in anni precedenti, di cui tre a distanza di un anno ed uno addirittura a distanza di otto.
Analisi biometrica
Il Piovanello è una specie monotipica che non mostra significanti variazioni geografiche nelle dimensioni. In tabella 5 sono raccolte le misure relative a peso, lunghezza alare, tarso e becco degli uccelli catturati.
Tabella 5. Peso (in g) e misure biometriche (in mm) dei Piovanelli, primavera 2002. Il sesso degli uccelli è stato determinato mediante la funzione discriminante proposta da Engelmoer (in Wymenga et al. 1990), analizzando le misure della lunghezza dell'ala e del becco. 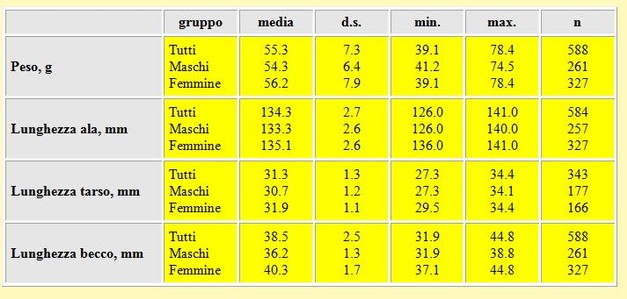
La differenza tra le medie delle lunghezze alari dei maschi e delle femmine è risultata altamente significativa (t-
Anche le differenze tra le medie delle lunghezze del tarso e del becco dei maschi e delle femmine sono risultate altamente (t-
Il peso medio calcolato per tutti gli uccelli è considerevolmente più basso rispetto ai valori rilevati in Camargue (Francia) in Maggio (media= 60.8 g d.s. 8.3 n= 66, Glutz et al.
Sesso ed età
Il sesso degli uccelli catturati è stato determinato mediante la funzione discriminante proposta da Engelmoer (in Wymenga et al. 1990), analizzando le misure della lunghezza dell'ala e del becco. Le catture hanno mostrato una deviazione della sex ratio da 1:1, con 56% di femmine (in totale sono stati catturati 261 maschi e 327 femmine). Questo risultato è in contrasto con quanto emerso dall'analisi dei dati raccolti nel periodo 1993-
L'età è stata determinata in base alle caratteristiche del piumaggio (Prater et al.
Analisi del grasso
I risultati dei pesi medi corrispondenti a ciascuna classe di grasso sono presentati nella tabella 6. È stata rilevata una significante relazione tra il peso degli uccelli e la quantità di grasso accumulata stimata visualmente: ad un più alto peso corrisponde una più alta classe di grasso, con una differenza tra le medie calcolate per ciascun peso per classe di grasso statisticamente significativa (t-
Tabella 6. Peso medio per ciascuna classe di grasso nei Piovanelli, primavera 2002.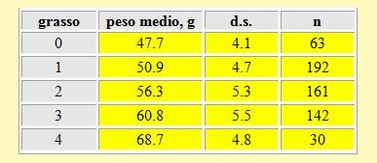
Variazione del peso nei ricatturati e lunghezza del periodo di sosta
Nel corso delle operazioni di cattura sono stati ricontrollati, a distanza di due/quattro giorni dall'inanellamento, soltanto quattro individui su 588 catture. È stata rilevata una sostanziale variazione di peso: i quattro uccelli hanno guadagnato peso, mostrando un incremento medio di 2.7 g/giorno (max 14.6 g, min 2.5 g), un valore molto alto se confrontato con quelli riportati in letteratura per aree dove avviene l'accumulo di grasso premigratorio (Meininger & Schekkerman 1994).
Sulla base dei giorni intercorsi tra cattura e ricattura è evidente che il periodo medio di sosta nell'area è molto breve, ma rispecchia quanto già accertato in altre aree, quali Grecia e Turchia.
Autonomia di volo
I Piovanelli che seguono la Mediterranean/Black Sea Flyway e migrano attraverso la Piana del Volturno utilizzano le aree umide della Grecia e dell'Ucraina meridionale come aree di sosta prima di arrivare nei territori riproduttivi; difatti, in primavera in queste zone umide sono stati ricatturati diversi uccelli inanellati negli anni precedenti.
Per confermare questa strategia di migrazione è stato calcolata l'autonomia di volo potenziale degli individui che lasciano la Piana del Volturno. Sono stati quindi utilizzati i seguenti valori di peso:
Peso magro (senza accumulo di grasso) = 47.7 g;
Peso alla partenza (con accumulo di grasso) = 68.7 g.
Con il modello matematico utilizzato (Davidson 1984) e con una velocità di volo di 41.2 km/h ipotizzata da Pennycuick (1989), è possibile stimare un'autonomia di volo potenziale di 1482 km per i Piovanelli che lasciano la Piana del Volturno. Ad una velocità di volo di circa 55 km/h accertata per i Limicoli e basata su osservazioni in campo (Zwarts et al. 1990) tale autonomia può arrivare a 1979 km. Queste distanze potrebbero anche essere sottostimate in quanto il valore del peso medio alla partenza potrebbe essere maggiore. In ogni caso, pur presentando una notevole differenza tra loro, consentirebbero ai Piovanelli che lasciano la Piana del Volturno di raggiungere le aree umide localizzate nella Grecia nord-
Piro piro boschereccio Tringa glareola
Questa specie nidifica nell'Europa nord-
Fenologia della migrazione
Le catture sono avvenute tra il 29 Marzo ed il 14 Maggio (pentadi 18-
Quest'anno, per la partecipazione al progetto "Tringa glareola 2000", 158 individui sono stati marcati anche con anelli colorati. Due inanellati dai centri d'inanellamento di Hiddensee (Germania) e Gdansk (Polonia) sono stati ricatturati, rispettivamente, il 12 e il 20 Aprile.
Analisi biometrica
In tabella 7 sono raccolte le misure relative a peso, lunghezza alare, terza remigante primaria, tarso, becco e nalospi degli uccelli catturati.
Tabella 7. Peso (in g) e misure biometriche (in mm) dei Piro piro boscherecci, primavera 2002.
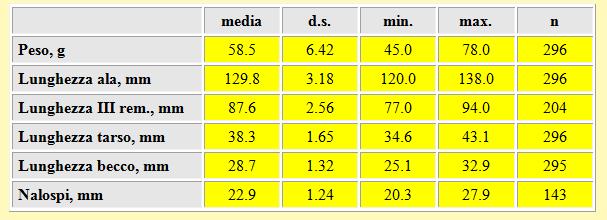
Le misure biometriche dell'ala, del becco e del tarso sono in accordo con quelle riportate da Cramp & Simmons (1983). Il peso medio è stato di 58.5 g (d.s.= 6.42, n= 296, min 45.0 g, max 78.0 g). La considerevole variazione che si rileva tra i due estremi può essere attribuita, per quanto riguarda i valori più bassi, alla cattura di uccelli che di recente hanno attraversato il Mediterraneo o, addirittura, sorvolato tutto o parte del Sahara, arrivando quindi nell'area di sosta con minime quantità di grasso e, per quanto riguarda i valori più elevati, alla cattura di uccelli che hanno già trascorso qualche giorno nell'area ed sono quindi riusciti a ripristinare le riserve di grasso. I pesi dei Piro piro boscherecci catturati in Camargue (Francia) sono risultati leggermente più alti rispetto a quelli rilevati nella Piana del Volturno: in Aprile è stato calcolato un peso medio di 62.6 g (d.s.= 7.1, n= 77) ed in Maggio di 61.6 g (d.s.= 6.4, n= 180) (Glutz et al. 1977); diversamente, quelli rilevati nella Grecia sud-
Sesso ed età
Non esistono differenze tra i sessi per quanto riguarda il piumaggio, ma vi sono delle modeste differenze nelle dimensioni, in quanto le femmine si presentano leggermente più grandi; in ogni caso, in campo non è possibile stabilire il sesso.
In base alla presenza di copritrici mediane interne di tipo giovanile, all'abrasione delle remiganti primarie e, in alcuni casi, al tipo di muta (Prater et al. 1977), 248 uccelli (84%) su 296 catturati sono stati identificati come individui nel secondo anno di età (nati nel 2001). Il campione raccolto presenta un rapporto giovani/adulti particolarmente elevato, pari a 5.2. Gli adulti (gli uccelli nati prima del 2001) tendono a passare nelle fasi iniziali della migrazione: nel primo quartile, cioè dal 29 Marzo al 20 Aprile, essi hanno rappresentato il 28% degli individui catturati; nelle fasi successive della migrazione, dal 21 Aprile in poi, questa percentuale si è ridotta al 12%.
Analisi del grasso
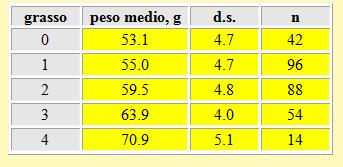
I risultati dei pesi medi corrispondenti a ciascuna classe di grasso sono presentati nella tabella 8. È possibile rilevare una significante relazione tra il peso degli uccelli e la quantità di grasso accumulata stimata visualmente: ad un più alto peso corrisponde una più alta classe di grasso, con una differenza tra le medie calcolate per ciascun peso per classe di grasso statisticamente significativa (t-
Tabella 8. Peso medio per ciascuna classe di grasso nei Piro piro boscherecci, primavera 2002.
Autonomia di volo
Nel corso della migrazione primaverile pochissime ricatture sono riportate in letteratura (Cramp & Simmons 1983). Per tentare di valutare le capacità di volo degli individui che lasciano la Piana del Volturno è stata calcolata l'autonomia di volo potenziale. Sono stati utilizzati i seguenti valori di peso:
Peso magro (senza accumulo di grasso) = 53.1 g;
Peso alla partenza (con accumulo di grasso) = 70.9 g.
Con il modello matematico utilizzato (Davidson 1984) e con una velocità di volo di 41.2 km/h ipotizzata da Pennycuick (1989), è possibile stimare un'autonomia di volo potenziale di 1191 km per i Piro piro boscherecci che lasciano la Piana del Volturno. Utilizzando una velocità di volo maggiore corrispondente a 55 km/h, determinata per i limicoli e basata su osservazioni in campo (Zwarts et al. 1990), l'autonomia di volo può arrivare a 1591 km.
Un'autonomia di volo potenziale di circa 1500 km consente ai Piro piro boscherecci che lasciano la Piana del Volturno di raggiungere direttamente l'Europa settentrionale con un unico volo senza soste intermedie. È inoltre possibile che individui con peso superiore a 71 g, dovuto all'accumulo di un'elevata quantità di grasso, possano compiere voli anche di 2500 km dirigendosi direttamente verso la penisola scandinava: nel 1998 un uccello catturato nell'area di studio e marcato con anelli colorati fu osservato in Finlandia dopo 12 giorni.
Considerazioni conclusive
I pochi ambienti umidi ancora presenti lungo il litorale sud-
Le profonde trasformazioni ambientali hanno notevolmente ridotto le possibilità di sosta per molte specie, ma ciò non significa che l'area non venga più attraversata da forti flussi migratori. Per la ridotta disponibilità di ambienti idonei, la presenza di un elevato numero di invasi artificiali costituisce una forte attrattiva per tutte le specie legate alle aree umide (in particolare Anatidi e Caradriformi) sia durante il periodo delle migrazioni che in inverno. Nel periodo primaverile persistono ancora ampie aree allagate in cui gli uccelli possono trovare facilmente notevoli quantità di cibo. La buona qualità dell'acqua consente difatti la presenza di piccoli crostacei (Gammarus sp.
Nella primavera del 2002 è stato realizzato il più alto numero di inanellamenti. La positiva esperienza legata ad uno sforzo di campionatura basato su almeno due uscite in campo per ogni pentade ed integrato da uscite continuative nel periodo più intenso della migrazione, ha consentito la raccolta di dati molto importanti relativi sia alle prime fasi della migrazione che a quelle terminali. E' comunque indubbio che il mese più fruttuoso in termini numerici è stato Maggio. L'assenza di attività nel periodo metà Febbraio-
L'area in cui è stata effettuata la ricerca non ha una valenza soltanto ornitologica. Nella primavera del 1996 è stata catturata una rara specie di pipistrello, un Myotis daubentoni
E' dunque evidente l'interesse naturalistico rivestito da quest'area e soprattutto da questi piccoli bacini artificiali per il ruolo che svolgono durante la migrazione. Va però sottolineato che la maggior parte dei terreni su cui sono stati scavati è di proprietà privata per cui la loro disponibilità è molto precaria ed è condizionata dall'attività venatoria. Molte vasche sono alimentate artificialmente pompando acqua dai canali vicini e solo alcune sono provviste di pozzi artesiani anch'essi scavati dai proprietari; pertanto, quando non sono più utilizzate, il livello dell'acqua si abbassa rapidamente e, già a metà maggio, molte sono asciutte o quasi. La presenza di aree allagate disponibili per gli uccelli durante la migrazione primaverile (ed in seguito anche per la nidificazione) è solo in parte dovuta alle precipitazioni atmosferiche; molto dipende dall'interesse dei proprietari a mantenere le vasche attive per potervi praticare la caccia durante la stagione venatoria.
E questo è l'aspetto gestionale più importante in quanto, fermo restando attualmente la chiusura della caccia al 31 Gennaio, occorre proporre delle soluzioni efficaci in grado di consentire la sosta ai grossi stormi di limicoli in migrazione. Una di queste potrebbe essere quella di consentire la caccia da appostamento fisso, come previsto dall'art. 5 della legge sulla caccia dell'11/02/92 n. 157, purché i titolari s'impegnino a mantenere adeguati livelli di acqua nelle vasche per tutto il periodo di migrazione e venga esercitata un'azione di controllo e di salvaguardia del territorio.
L'attuale livello di conoscenze sulle variazioni di densità e sulle strategie di migrazione adottate dalle varie popolazioni che utilizzano la "Mediterranean/Black Sea Flyway" durante la migrazione di ritorno verso le aree di nidificazione, è decisamente ancora inferiore rispetto a quanto è stato invece accertato dalle ricerche svolte sulle popolazioni che seguono la "East Atlantic Flyway". I dati raccolti nel corso di questo primo anno del progetto "Monitoraggio delle popolazioni di Limicoli nella Piana del Volturno (Caserta) durante la migrazione primaverile", e naturalmente anche quelli degli anni precedenti, consentono di investigare sia gli aspetti generali che di assoluto dettaglio. Essi confermano gli ottimi risultati scientifici dell'attività svolta ai fini del monitoraggio dell'avifauna selvatica in quanto forniscono preziose informazioni in relazione al numero assoluto degli animali in migrazione, alle variazioni negli anni, al rilevamento di specie rare, all'analisi della fenologia delle specie migratorie.
Sarebbe comunque opportuno integrare i dati raccolti con un'indagine sulle disponibilità alimentari dell'area, così come sarebbe di fondamentale importanza anche la collaborazione con altre stazioni europee sia nella raccolta che nell'analisi dei dati, in modo da ottenere un quadro più completo sulle strategie migratorie adottate e sulle rotte seguite durante la migrazione pre-
Una parte dei dati fin'ora acquisiti è già stata pubblicata su riviste nazionali ed internazionali; in ogni caso le informazioni raccolte possono essere utilizzate dalla pubblica amministrazione come un'utile guida per la futura protezione e gestione dell'ambiente, per tentare di avviare un miglioramento delle condizioni ambientali dell'intera area e per giustificare la conservazione di una catena di aree umide di limitata estensione in grado di consentire il passaggio e la sosta degli uccelli migratori. L'attività svolta può inoltre servire a sensibilizzare maggiormente le associazioni venatorie locali per quanto riguarda i problemi connessi con la caccia, impegnandole più direttamente in un'attività di protezione e salvaguardia delle ultime zone umide.
Ringraziamenti
Un particolare ringraziamento va ad Eduardo Paparo, Delegato Provinciale LIPU di Caserta, a Tonino D'Angelo, Presidente Regionale UNAVI ed a Riccardo Ventre, Presidente dell'A.T.C. e della Provincia di Caserta, membri del Comitato di Gestione A.T.C. Caserta, per l'interesse dimostrato e l'impegno profuso nel portare avanti questa ricerca. Un doveroso ringraziamento va anche a Costantino ed Andrea Tamburrino per la loro sempre squisita ospitalità e disponibilità ed a John Glazebrook, Oreste Martinelli, Alan Miller, Olena Tryasko e Anna Vannucchi per l'indispensabile aiuto in campo. Si ringraziano infine tutti i proprietari delle vasche che hanno gentilmente consentito l'accesso sui loro terreni permettendo di portare a termine con successo questa prima stagione di studio. La ricerca è stata realizzata con il finanziamento del Comitato di Gestione dell'A.T.C. di Caserta.
Riassunto
Il Gruppo Inanellamento Limicoli (G.I.L., Napoli) ha avviato dal 1991 un programma di studio a lungo termine basato su campagne di inanellamento e osservazioni allo scopo di effettuare un monitoraggio costante dei movimenti migratori dei Caradriformi nella Piana del Volturno (Caserta).
Per la ridotta disponibilità di ambienti idonei, la presenza di un elevato numero di invasi artificiali utilizzati per scopi venatori costituisce una forte attrattiva per tutte le specie legate alle aree umide (in particolare Anatidi e Caradriformi) sia durante il periodo delle migrazioni che in inverno. Frequentemente nel corso della migrazione primaverile sono stati osservati stormi anche di 2000/3000 Limicoli alimentarsi attivamente in vasche di modesta estensione.
Viene qui presentata un'analisi preliminare dei dati raccolti nel corso della migrazione primaverile del 2002; da quest'anno la ricerca viene condotta per conto del Comitato di Gestione dell'Ambito Territoriale Caccia di Caserta che si propone di effettuare una ricognizione delle risorse ambientali e della consistenza faunistica della provincia.
La positiva esperienza legata ad uno sforzo di campionatura basato su almeno due uscite in campo per ogni pentade ed integrato da uscite continuative nel periodo più intenso della migrazione, ha consentito la raccolta di dati molto importanti relativi sia alle prime fasi della migrazione che a quelle terminali. In totale sono stati inanellati 1.559 uccelli appartenenti a 27 specie. Il Piovanello (Calidris ferruginea) ha caratterizzato il 2002, in quanto ha costituito il 37% dell'intera popolazione inanellata. È importante rimarcare anche la cattura di alcune specie di notevole importanza per l'intero territorio nazionale e regionale, incluse nell'Allegato I della Direttiva 79/409/CEE ed in particolare il Piro piro boschereccio (Tringa glareola), il Cavaliere d'Italia (Himantopus himantopus), il Combattente (Philomachus pugnax), il Mignattino (Chlidonias niger) ed il Tarabusino (Ixobrychus minutus).
Nel periodo di studio la migrazione è stata caratterizzata dalla presenza di tre diverse ondate: la prima ad inizio Aprile (pentade 20), determinata per lo più dal passaggio di Combattenti, soprattutto femmine; quindi i numeri sono andati diminuendo per poi risalire nuovamente verso la fine di Aprile (pentade 23) quando le catture sono aumentate con un andamento abbastanza regolare fino ad un forte passaggio agli inizi di Maggio (pentade 25). Una terza ondata migratoria, dovuta ad un intenso transito di Piovanelli e Gambecchi, si è verificata nella seconda metà di Maggio.
Numerose anche le osservazioni di specie incluse nell'Allegato I: Spatola (Platalea leucorodia), Airone rosso (Ardea purpurea), Pernice di mare (Glareola pratincola), Avocetta (Recurvirostra avosetta), Gabbiano roseo (Larus genei), Sterna maggiore (Sterna caspia), Beccapesci (Sterna sandvicensis), Sterna comune (Sterna hirundo) e Mignattino piombato (Chlidonias hybridus), per le quali sono stati accertati regolari movimenti migratori.
La Piana del Volturno rappresenta un'importante zona di sosta lungo la Mediterranean/Black Sea Flyway, in quanto durante la migrazione pre-
Per tutte e tre le specie è stata riscontrata una significante relazione tra il peso medio e la quantità di grasso stimata visualmente: le medie più alte corrispondono evidentemente alle classi di grasso maggiori, con differenze statisticamente significanti tra le medie di ciascun peso per classe di grasso. Sulla base dei giorni intercorsi tra cattura e ricattura è emerso che il periodo medio di sosta nell'area è molto breve, ma rispecchia comunque quanto già accertato in altre aree, quali Grecia e Turchia.
È stato anche accertato che Gambecchi e Piovanelli che lasciano la Piana del Volturno posseggono un'autonomia di volo potenziale che consente loro di attraversare l'Adriatico per raggiungere altre aree umide quali, ad esempio, quelle localizzate tra il delta del Nestos ed il delta dell'Evros lungo le coste nord-
Il numero dei Piovanelli inanellati nel corso di questo primo anno del progetto è risultato il più alto rispetto a quelli realizzati negli anni precedenti. Queste fluttuazioni rientrano comunque nel normale andamento della fenologia della migrazione in quanto, da un anno all'altro, è possibile che si verifichino delle grosse variazioni nei passaggi dei migratori. Come già è stato evidenziato dai dati raccolti negli anni precedenti, i maschi hanno mostrato la tendenza a precedere le femmine:
Allo scopo di incrementare le segnalazioni, è proseguito il marcaggio di individui di Piro piro boschereccio con una combinazione di quattro anelli colorati che consentono di risalire alla località di cattura, all'anno ed alla pentade in cui è avvenuto l'inanellamento. Questa attività viene svolta nell'ambito della partecipazione al progetto "Tringa glareola 2000", coordinato dal Kuling Group (Polonia) e finalizzato allo studio della migrazione di questa specie nel Paleartico.
I Piro piro boscherecci che lasciano la Piana del Volturno hanno presentato un'autonomia di volo potenziale di circa 1500 km che consente loro di raggiungere direttamente l'Europa settentrionale con un unico volo senza soste intermedie. È inoltre possibile che individui con elevate quantità di grasso possano compiere voli anche di 2500 km dirigendosi direttamente verso la penisola scandinava.
Dai dati raccolti è evidente l'interesse naturalistico rivestito dalla Piana del Volturno e soprattutto da questi piccoli bacini artificiali per il ruolo che svolgono durante la migrazione. Va però sottolineato che la maggior parte dei terreni su cui sono stati scavati è di proprietà privata per cui la loro disponibilità è molto precaria ed è condizionata dall'attività venatoria. La presenza di aree allagate disponibili per gli uccelli durante la migrazione primaverile (ed in seguito anche per la nidificazione) è solo in parte dovuta alle precipitazioni atmosferiche; molto dipende dall'interesse dei proprietari a mantenere le vasche attive per potervi praticare la caccia durante la stagione venatoria. E questo è l'aspetto gestionale più importante in quanto, fermo restando attualmente la chiusura della caccia al 31 Gennaio, occorre proporre delle soluzioni efficaci in grado di migliorare le condizioni ambientali dell'intera area ed assicurare la conservazione di una catena di aree umide di limitata estensione in grado di consentire il passaggio e la sosta degli uccelli migratori.
Abstract
Numbers of migrating waders on the Volturno Plain (Caserta) assessed by standardised monitoring during spring migration 2002
The waders are a common migrant in southern Italy but its migration dynamics and weight changes are undescribed and ringing of species in this family has always been of an occasional manner. Little is known about wader migrating routes in the Mediterranean area, the utilization of food resources and the changes in body mass during periods at stopover site in Campania.
A 3-
-
-
-
-
The work is carried out on the Volturno Plain (Caserta) in the north west of Campania region along the south side of Volturno river. Originally, a marsh enlarged on the whole plain formed by the ancient river Clanio. The drainage of the zone began about in 1539 and it was continued until the end of the past century; also today there are some works of canalization, so the plain appears to be crossed by a large number of canals, named "Regi Lagni". Consequently, even if the cultivation of the area has developed during the last years, otherwise the place has lost its original naturalistic interest.
The area between Patria Lake and the mouth of Volturno, for 3-
We mist-
Approximately every day all waders in the study area are counted by car and on foot. The counts are carried out in the afternoon, when the birds are feeding and roosting.
General information on the Little Stints, Curlew Sandpipers and Wood Sandpipers, the timing and intensity of migration, morphology of the migrants and details about supposed migration routes are given. Some details about the timing of passage of males and females are given and some morphometric data and weight changes are examined; birds show a significant mass increase during the study season and a short stopover duration was ascertained.
Results can be used to justify the necessity of the maintenance of a chain of small wetland areas to provide safe passage for migratory birds. Results of the project will be presented to the public administration and the media as guide for future nature protection.
Bibliografia
Akriotis, T. (1991) Weight changes in the Wood Sandpiper Tringa glareola in south-
Berthold, P. (1973) Proposals for the standardization of the presentation of annual events, especially of migrating data. Auspicium
Berthold, P. & Friedrich, W. (1979) Die Federlänge: Ein neues nützliches Flügelmass. Vogelwarte
Chernichko, I.I., Chernichko, R., Diadicheva, E., Van der Have, T.M. & Van de Sant, S. (1993) Biometry of waders in the Sivash, Ukraine, spring 1992. In: Waterbirds in the Sivash, Ukraine, spring 1992. T.M.van der Have, S. van de Sant, Y. Verkuil and J. van der Winden (eds.). WIWO-
Cramp, S. & Simmons, K.E.L. (1983) The Handbook of the Birds of the Western Palearctic.
Davidson, N.C. (1984) How valid are flight range estimates for waders? Ringing & Migration, 5: 49-
Glutz von Blotzheim, U.N., Bauer, K.M. & Bezzel, E. (1975) Handbuch der Voegel Mitteleuropas.
Glutz von Blotzheim, U.N., Bauer, K.M. & Bezzel, E. (1977) Handbuch der Voegel Mitteleuropas. Vol. 7. Akademische. Verlagsgellsschaft, Wiesbaden.
Hötker, H., Lebedeva, E., Tomkovich, P.S., Gromadzka, J., Davidson, N.C., Evans, J., Stroud, D.A. & West, R.B. (eds) (1998) Migration and international conservation of waders. Research and conservation on north Asian, African and European flyways. International Wader Studies 10.
Karr, J.R. (1981) Surveying birds in the tropics. Studies in Avian Biology N° 6: 548-
Meininger, P.L. (1990) Birds of the wetlands in north-
Meininger, P.L. & Schekkerman, H. (1994) Measurements, moult and mass of waders captured in Egypt, winter and spring 1990. In: Meininger, P.L. & Atta G.A.M. (eds.). Ornithological studies in Egyptian wetlands 1989/90. WIWO-
Moschetti, G. & Scebba, S. (1999) Migration pattern and fat reserves of Curlew Sandpiper Calidris ferruginea
Pennycuick, C.J. (1989) Bird flight performance. A practical calculation manual. Oxford Science Publications, Oxford.
Prater, A.J., Marchant, J.H. & Vuorinen, J. (1977) Guide to the Identification and Ageing of Holarctic Waders. BTO Guide 17. Tring, Herts.
Russo, D., Picariello, O. e Moschetti, G. (1995) Presenza del Vespertilio di Daubenton Myotis daubentoni (Kuhl, 1819) in Campania (Mammalia, Chiroptera). Ucc. d'Italia, 20: 140-
Scebba, S. (1993) Gli uccelli della Campania
Scebba, S., Moschetti, G., Lancini, M. e Esposito, R. (1993) Dati preliminari sulla migrazione dei Limicoli in Campania. Atti VII Conv. ital. Ornit., Urbino. In: Ric. Biol. Selvaggina XXII (1995): 473-
Scebba, S., Moschetti, G. e Taglialatela, D. (1994) Importanza delle vasche artificiali nella Piana del Volturno (Caserta) per la migrazione dei Caradriformi. In: Fraissinet M., Coppola D., Del Gaizo S., Grotta M. e Mastronardi D. (red.) 1995. Atti Convegno Nazionale "L'avifauna degli ecosistemi di origine antropica: zone umide artificiali, coltivi, aree urbane".
Scebba, S. e Moschetti, G. (1995) Prima nidificazione accertata di Pernice di mare Glareola pratincola
Scebba, S. & Moschetti, G. (1996) Migration pattern and weight change of Wood Sandpiper Tringa glareola
Scebba, S. & Moschetti, G. (in press) Migration pattern and fat reserves of Curlew Sandpiper Calidris ferruginea
Spiekman, H. (1990) Waterbird trapping. In: Meininger, P.L. (ed.). Birds of the wetlands in north-
Spiekman, H. (1992) Results of wader ringing activities in Tunisia 1962-
Svensson, L. (1992) Identification guide to European passerines.
Wylson, J.R., Czajkowski, M.A., Pienkowski, M.W. (1980) The migration through Europe and wintering in West Africa of the Curlew Sandpiper. Wildfowl, 31: 107-
Wymenga, E., Engelmoer, M., Smit, C.J. & Van Spanje, T.M. (1990) Geographical breeding origin and migration of waders wintering in West Africa. Ardea
Zwarts, L., Ens, B.J., Kersten, M. & Piersma, T. (1990) Moult, mass and flight range of waders ready to take off for long-
Pubblicazione n° 43 del Gruppo Inanellamento Limicoli, Napoli